Потенциал действия (ПД) — это кратковременные амплитудные изменения мембранного потенциала покоя (МПС), возникающие при возбуждении живой клетки. По сути это электрический разряд — быстрая кратковременное изменение потенциала на небольшом участке мембраны возбудимой клетки (нейрона или мышечного волокна), в результате которого внешняя поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны. Потенциал действия является физической основой нервного или мышечного импульса, который играет сигнальную (регуляторную) роль.
Общая характеристика
Потенциалы действия могут отличаться по своим параметрам в зависимости от типа клетки и даже на разных участках мембраны одной и той же клетки. Наиболее характерный пример различий: потенциал действия сердечной мышцы и потенциал действия большинства нейронов. Все же, в основе любого потенциала действия лежат следующие явления:
- «Мембрана живой клетки поляризована» — ее внутренняя поверхность заряжена отрицательно по отношению к наружной благодаря тому, что в растворе у ее внешней поверхности находится большее количество положительно заряженных частиц (катионов), а у внутренней поверхности — большее количество отрицательно заряженных частиц (анионов).
- «Мембрана имеет избирательную проницаемость ‘- ее проницаемость для различных частиц (атомов или молекул) зависит от их размеров, электрического заряда и химических свойств.
- «Мембрана возбудимой клетки способна быстро менять свою проницаемость ‘для определенного вида катионов, вызывая переход положительного заряда с внешней стороны на внутреннюю
Первые два свойства характерны для всех живых клеток. Третья же является особенностью клеток возбудимых тканей и причиной, по которой их мембраны способны генерировать и проводить потенциалы действия.
Основной математической моделью, описывающей генерацию и передачу потенциала действия, является модель Ходжкина-Хаксли.
Деполяризация и реполяризация
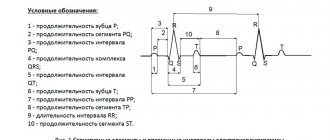
В разделе «Основные принципы электрокардиографии и нарушения» рассматривается общее понятие «электрическое возбуждение», которое означает распространение электрических импульсов по предсердиям и желудочкам. Точное название электрического возбуждения, или активации сердца, – деполяризация. Возврат кардиомиоцитов к состоянию расслабления после возбуждения (деполяризации) – реполяризация. Эти термины подчёркивают, что в состоянии покоя клетки миокарда предсердий и желудочков поляризованы (их поверхность электрически заряжена). На рисунке 2-1, А изображено состояние поляризации нормальной мышечной клетки предсердий или желудочков
Рис. 2-1. Процессы деполяризации и реполяризации А – мышечная клетка сердца в состоянии покоя поляризована, т.е. наружная поверхность клетки заряжена положительно, а внутренняя – отрицательно; Б — при возбуждении клетки (S) происходит её деполяризация (возбуждённый участок электроотрицателен по отношению к соседним участкам); В – полностью деполяризованная клетка заряжена положительно внутри и отрицательно – снаружи; Г – реполяризация происходит, когда клетка возвращается из состояния возбуждения в состояние покоя. Направление деполяризации и реполяризации указано стрелками. Деполяризации (возбуждению) предсердий на ЭКГ соответствует зубец Р, а деполяризации желудочков – комплекс QRS. Реполяризации желудочков соответствует комплекс ST-T.
Внешняя сторона клетки в состоянии покоя заряжена положительно, а внутренняя сторона – отрицательно [около -90 мВ (милливольт)]. Поляризация мембраны обусловлена разницей концентраций ионов внутри и вне клетки.
При возбуждении мышечной клетки сердца происходит её деполяризация. В результате внешняя сторона клетки в области возбуждения становится отрицательной, а внутренняя сторона – положительной. Возникает разница электрического напряжения на внешней поверхности мембраны между деполяризованным участком в состоянии возбуждения и невозбуждённым поляризованным участком см. рис. 2-1, Б. Затем возникает небольшой электрический ток, который распространяется вдоль клетки до её полной деполяризации см. рис. 2-1, В.
Направление деполяризации показано стрелкой см. рис. 2-1, Б. Деполяризация и реполяризация отдельных мышечных клеток (волокон) происходит в одном направлении. Однако во всём миокарде деполяризация идёт от внутреннего слоя (эндокардиального) к наиболее удалённому слою (эпикардиальному), а реполяризация – в противоположном направлении. Механизм этого различия до конца не ясен.
| Примечание: Деполяризация миокарда желудочков происходит от эндокарда к эпикарду, а реполяризация – от эпикарда к эндокарду. Это обусловлено тем, что длительность ТМПД в субэпикардиальных отделах желудочков на 0,03-0.04 с меньше, чем в субэндокардиальных отделах, и процесс реполяризации раньше начнётся именно под эпикардом. | |
Деполяризующий электрический ток регистрируют на электрокардиограмме в виде зубца Р (возбуждение и деполяризация предсердий) и комплекса QRS (возбуждение и деполяризация желудочков).
Через некоторое время полностью охваченная возбуждением деполяризованная клетка начинает возвращаться к состоянию покоя. Этот процесс называют реполяризацией. Небольшой участок на внешней стороне клетки вновь приобретает положительный заряд см. рис. 2-1, Г, затем процесс распространяется вдоль клетки до её полной реполяризации. Реполяризации желудочков на электрокардиограмме соответствуют сегмент ST, зубцы и (реполяризация предсердий обычно скрыта потенциалами желудочков).
На электрокардиограмме отражена электрическая активность всех клеток предсердий и желудочков, а не отдельных клеток. В сердце деполяризация и реполяризация обычно синхронизированы, поэтому на электрокардиограмме можно записать эти электрические потоки в виде определённых зубцов (зубцы P, T, U, комплекс QRS, сегмент ST).
| Любая электрокардиограмма – и нормальная, и патологически изменённая – отражает два основных процесса: деполяризацию – распространение импульса по миокарду – и реполяризацию – возвращение возбуждённого миокарда в состояние покоя. | |
Фазы
Можно четко выделить пять фаз развития ПД:
Нарастание (деполяризация)
Возникновение потенциала действия (ПД) связано с увеличением проницаемости мембраны для ионов натрия (в 20 раз по сравнению с проницаемостью для К +, и в 500 раз по сравнению с исходной проницаемостью Na +) и последующим усилением диффузии этих ионов по концентрационном градиенту внутрь клетки, приводит к изменению (уменьшение) мембранного потенциала. Уменьшение мембранного потенциала приводит к увеличению проницаемости мембраны для натрия путем открытия потенциал-зависимых натриевых каналов, а увеличение проницаемости сопровождается усилением диффузии натрия в цитоплазму, что вызывает еще более значительную деполяризацию мембраны. Благодаря наличию положительной обратной связи деполяризация мембраны при возбуждении происходит с ускорением и поток ионов натрия в клетку все время растет. Интенсивность же потока ионов калия, направленного из клетки наружу, в первые моменты возбуждения остается в начале. Усиленный поток положительно заряженных ионов натрия внутрь клетки вызывает сначала исчезновение избыточного отрицательного заряда на внутренней поверхности мембраны, а затем приводит к перезарядки мембраны. Поступления ионов натрия происходит до тех пор, пока внутренняя поверхность мембраны не приобретет положительный заряд, достаточный для уравновешивания градиента концентрации натрия и прекращение его дальнейшего перехода внутрь клетки. Натриевый возникновения ПД подтверждают опыты с изменением внешней и внутренней концентрации этого иона. Было показано, что десятикратном изменении концентрации ионов натрия во внешнем или внутреннем среде клетки, соответствует изменение ПД на 58 мВ. При полном удалении ионов натрия из окружающей клетку жидкости ПД ни возникал. Таким образом, установлено, что ПД возникает в результате избыточной, по сравнению с покоем, диффузии ионов натрия из окружающей жидкости внутрь клетки. Период, в течение которого проницаемость мембраны для ионов натрия при открытии натриевых каналов растет, является небольшим (0,5-1 мс) вслед за этим наблюдается повышение проницаемости мембраны для ионов калия благодаря открытию потенциал-зависимых калиевых каналов, и, следовательно, усиление диффузии этих ионов из клетки наружу.
Принцип «все или ничего» Согласно закону «все-или-ничего» мембрана клетки возбудимой ткани или не отвечает стимул совсем, или отвечает с максимально возможной для нее на данный момент силой. Действие раздражителя обычно приводит к локальной деполяризации мембраны. Это вызывает открытие натриевых каналов, которые чувствительны к изменениям потенциала, а через это — увеличивает натриевую проводимость, что приводит к еще большей деполяризации. Существование такой обратной связи обеспечивает регенеративную (возобновляемую) деполяризацию клеточной мембраны. Величина потенциала действия зависит от силы раздражителя, а сам он возникает только в том случае, когда деполяризация превышает некоторый определенный для каждой клетки предельный уровень. Это явление получило название «все или ничего». Однако, если деполяризация составляет 50-75% от предельной величины, то в клетке может возникнуть локальный ответ, амплитуда которой значительно ниже амплитуду потенциала действия. Отсутствие потенциала действия при пидграничному уровне деполяризации объясняется тем, что при этом недостаточно увеличивается натриевая проницаемость, чтобы вызвать регенеративную деполяризацию. Уровень деполяризации, который возникает при этом, не вызывает открытие новых натриевых каналов, поэтому натриевая проводимость быстро уменьшается, и в клетке снова устанавливается потенциал покоя.
Овершут
Деполяризация мембраны приводит к реверсии мембранного потенциала (МП становится положительным). В фазу овершута Na + -ток начинает стремительно снижаться, что связано с инактивацией потенциал-зависимых Na + -каналов (время открытого состояния — судьбы миллисекунды) и исчезновением электрохимического градиента Na +.
Рефрактерность Одним из последствий исчезновения градиента Na + является рефрактерность мембраны — временная неспособность отвечать на раздражитель. Если раздражитель возникает сразу после прохождения потенциала действия, то возбудимость не возникнет ни при силе раздражителя на уровне порога, ни при значительно более сильное раздражителю. Такое положение полной невозбудимости называется абсолютным рефрактерным периодом. За ним следует относительный рефрактерный период, когда надпороговый раздражитель может вызвать потенциал действия со значительно меньшей амплитудой чем в норме. Потенциал действия привычной амплитуды при действии порогового раздражителя можно вызвать только после нескольких миллисекунд после предварительного потенциала действия. Абсолютный рефрактерный период ограничивает максимальную частоту генерации потенциалов действия.
Реполяризация
Увеличение ионного потока калия, направленного из клетки наружу, приводит к уменьшению мембранного потенциала, в свою очередь обусловливает уменьшение проницаемости мембраны для ионов натрия, что, как указывалось, является функцией мембранного потенциала. Таким образом, второй этап характеризуется тем, что поток ионов калия из клетки наружу растет, а встречный поток ионов натрия уменьшается. Такая реполяризация мембраны продолжается, пока не произойдет восстановление потенциала покоя — реполяризация мембраны. После этого проницаемость для ионов калия также падает до исходной величины. Внешняя поверхность мембраны за счет положительно заряженных ионов калия, вышедших в среду, вновь приобретает положительный потенциала относительно внутреннего.
Следовая деполяризация и гиперполяризация
В конечной фазе происходит замедление восстановления мембранного потенциала покоя, и при этом регистрируются следовые реакции в виде следовой деполяризации и гиперполяризации, обусловлены медленным восстановлением исходной проницаемости для ионов К +.
Соотношение фаз потенциала действия и возбудимости.
Уровень возбудимости клетки зависит от фазы ПД. В фазу локального ответа возбудимость возрастает. Это фазу возбудимости называют латентным дополнением.
В фазу реполяризации ПД, когда открываются все натриевые каналы и ионы натрия лавинообразно устремляются в клетку, никакой даже сверхсильный раздражитель не может стимулировать этот процесс. Поэтому фазе деполяризации соответствует фаза полной невозбудимости или абсолютной рефрактерности.
В фазе реполяризации все большая часть натриевых каналов закрывается. Однако они могут вновь открываться при действии сверхпорогового раздражителя. Т.е. возбудимость начинает вновь повышаться. Этому соответствует фаза относительной невозбудимости или относительной рефрактерности.
Во время следовой деполяризации МП находится у критического уровня, поэтому даже допороговые стимулы могут вызвать возбуждение клетки. Следовательно в этот момент ее возбудимость повышена. Эта фаза называется фазой экзальтации или супернормальной возбудимости.
В момент следовой гиперполяризации МП выше исходного уровня, т.е. дальше КУД и ее возбудимость снижена. Она находится в фазе субнормальной возбудимости. Рис. Следует отметить, что явление аккомодации также связано с изменением проводимости ионных каналов. Если деполяризующий ток нарастает медленно, то это приводит к частичной инактивации натриевых, и активации калиевых каналов. Поэтому развития ПД не происходит.
Мембранный потенциал (МП) – разность потенциалов между наружной и внутренней стороной мембраны в состоянии физиологического покоя.
Причины возникновения МП:
1. неодинаковое распределение ионов по обе стороны мембраны: внутри — больше К+, снаружи – его мало, но больше Nа+ и Cl. такое распределение ионов называется ионной ассиметрией.
2. избирательная проницаемость мембраны для ионов. В состоянии покоя мембрана неодинакова проницаема.
За счет этих факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта в результате разности концентрации ионов.
Ионы К выходят из клетки и увеличивают положительный заряд на наружной поверхности мембраны. Сl — пассивно переходит во внутрь клетки, что приводит к повышению положительного заряда на наружной поверхности мембраны. Nа накапливается на наружной поверхности мембраны и увеличивает «+» заряд. Органические соединения остаются внутри клетки.
В результате такого движения наружная поверхность мембраны «+» заряжена, а внутренняя «-». Внутренняя поверхность может быть «-» заряжена, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние называется поляризацией.
Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов, т.е. пока не наступит электрохимическое равновесие.
Момент равновесия зависит от двух сил:
1. сила диффузии
2. Сила электрохимического взаимодействия.
Значение электрохимического равновесия:
3. поддержание ионной асимметрии
4. поддержание величины мембранного потенциала на постоянном уровне.
Возникновение МП при участи двух сил называют концентрационно-электрохимическим.
Для поддержания ионной симметрии электрохимического равновесия в клетке имеется Nа-К насос. В клеточной мембране имеется система переносчиков, каждый из которых связывает 3Na, которые находятся снаружи, а с внутренней стороны переносчик связывает 2К и переносит внутрь клетки. При этом расходуется 1 молекула АТФ.
Работа Nа-К насоса обеспечивает:
1. высокую концентрацию К внутри клетки, т.е. постоянную величину потенциала покоя
2. низкую концентрацию Nа внутри клетки, т.е. сохраняется нормальная осмомолярность, объем клетки, создает базу для генерации ПД.
3. стабильный концентрационный градиент Nа, способствуя транспорту аминокислот и сахаров.
МП в норме: для гладких мышц -30 – (-70) мВ, для нерва -50 – (-70) мВ, для миокарда -60 – (-90) мВ.
Потенциал действия (ПД) – сдвиг потенциала покоя, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой мембраны.
При действии порогового и сверхпорогового раздражителей изменяется проницаемость клеточной мембраны для ионов. Для Nа увеличивается в 450 раз и градиент нарастает быстро. Для К увеличивается в 10-15 раз и градиент развивается медленно. В результате движение Nа происходит внутрь клетки, К двигается из клетки, что приводит к перезарядке клеточной мембраны.
Фазы:
0. Локальный ответ (местная деполяризация), предшествующий развитию ПД.
1. Фаза деполяризации. Во время этой фазы МП быстро уменьшается и достигает нулевого уровня. Уровень деполяризации растет выше 0. Поэтому мембрана приобретает противоположный заряд — внутри она становится положительной, а снаружи отрицательной. Явление смены заряда мембраны называется реверсией мембранного потенциала. Продолжительность этой фазы у нервных и мышечных клеток 1-2 мсек.
2. Фаза реполяризации. Она начинается при достижении определенного уровня МП (примерно +20 мВ). Мембранный потенциал начинает быстро возвращаться к потенциалу покоя. Длительность фазы 3-5 мсек.
3. Фаза следовой деполяризации или следового отрицательного потенциала. Период, когда возвращение МП к потенциалу покоя временно задерживается. Он длится 15-30 мсек.
4. Фаза следовой гиперполяризации или следового положительного потенциала. В эту фазу, МП на некоторое время становится выше исходного уровня ПП. Ее длительность 250-300 мсек.
Возникновение ПД обусловлено изменением ионной проницаемости мембраны при возбуждении. В период локального ответа открываются медленные натриевые каналы, а быстрые остаются закрытыми, возникает временная самопроизвольная деполяризация. Когда МП достигает критического уровня, закрытые активационные ворота натриевых каналов открываются и ионы натрия лавинообразно устремляются в клетку, вызывая нарастающую деполяризацию. В эту фазу открываются и быстрые и медленные натриевые каналы. Т.е. натриевая проницаемость мембраны резко возрастает. Причем от чувствительности активационных зависит величина критического уровня деполяризации, чем она выше, тем ниже КУД и наоборот.
Когда величина деполяризация приближается к равновесному потенциалу для ионов натрия (+20 мВ), сила концентрационного градиента натрия значительно уменьшается. Одновременно начинается процесс инактивации быстрых натриевых каналов и снижения натриевой проводимости мембраны. Деполяризация прекращается. Резко усиливается выход ионов калия, т.е. калиевый выходящий ток. В некоторых клетках это происходит из-за активации специальных каналов калиевого выходящего тока. Этот ток, направленный из клетки, служит для быстрого смещения МП к уровню потенциала покоя. Т.е. начинается фаза реполяризации. Возрастание МП приводит к закрыванию и активационных ворот натриевых каналов, что еще больше снижает натриевую проницаемость мембраны и ускоряет реполяризацию.
Возникновение фазы следовой деполяризации объясняется тем, что небольшая часть медленных натриевых каналов остается открытой.
Следовая гиперполяризация связана с повышенной, после ПД, калиевой проводимостью мембраны и тем, что более активно работает натрий-калиевый насос, выносящий вошедшие в клетку во время ПД ионы натрия.
Распространение
Распространение в немиелинизированные волокне
В немиелинизированные (без`мякотному) нервном волокне ПД распространяется от точки к точке, поскольку возбуждение можно зарегистрировать как такое, что постепенно «бежит» по всему волокну от места своего возникновения. Ионы натрия, входящих внутрь возбуждении участка, служат источником электрического тока для возникновения ПД в прилегающих участках. В этом случае импульс возникает между деполяризована участком мембраны и ее невозбужденном участком. Разность потенциалов здесь во много раз выше, чем необходимо для того, чтобы деполяризация мембраны достигла предельного уровня. Скорость распространения импульса в таких волокнах 0,5-2 м / с
Распространение в миелинизированные волокне
Нервные отростки большинства соматических нервов миелинизированные. Только очень незначительные их участки, так называемые перехвата узла (перехват Ранвье), покрытые обычной клеточной мембраной. Такие нервные волокна характеризуются тем, что на мембране только в перехватах размещении потенциал-зависимые ионные каналы. Кроме того, эта оболочка повышает электрическое сопротивление мембраны. Поэтому при сдвиге мембранного потенциала ток проходит через мембрану перехватывающих участка, то есть прыжками (сальтаторно) от одного перехвата к другому, что позволяет увеличить скорость проведения нервного импульса, которая составляет от 5 до 120 м / с. Причем потенциал действия, который возник в одном из перехватов Ранвье, вызывает потенциалы действия в соседних перехвата за счет возникновения электрического поля, которое вызывает начальную деполяризацию в этих перехватов. Параметры ЭДС поля и дистанция его эффективного действия зависят от кабельных свойств аксона.
Типы нервных волокон, скорость проведения импульса, в зависимости от миелинизации
| Тип | Диаметр (мкм) | Миелинизация | Скорость проведения (м / с) | Функциональное назначение |
| А alpha | 12-20 | сильная | 70-120 | Подвижные волокна соматической НС; чувствительные волокна проприорецепторов |
| А beta | 5-12 | сильная | 30-70 | Чувствительные волокна рецепторов кожи |
| А gamma | 3-16 | сильная | 15-30 | Чувствительные волокна проприорецепторов |
| А delta | 2-5 | сильная | 12-30 | Чувствительные волокна терморецепторов, ноцицепторов |
| В | 1-3 | слабая | 3-15 | Преганглионарные волокна симпатической НС |
| С | 0,3-1,3 | отсутствует | 0,5-2,3 | Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов некоторых механорецепторов |
2_3 Потенциал действия и нервный импульс
Нервный импульс
Разбираясь в нервных импульсах, мы будем иметь в виду нервное возбуждение, бегущее (=распространяющееся) по мебране нейрона. Строго говоря, движущееся по нейронам и нервам возбуждение представляет собой нервные импульсы, а не потенциалы действия, хотя в физиологической литературе два этих понятия обычно используют как синонимы.
Для того чтобы произвести нервный импульс, нейрон сначала должен создать состояние готовности (потенциал покоя), затем предготовности (локальный потенциал), и, наконец, при достижении порогового уровня локального потенциала (критического уровня деполяризации) — породить бегущий по мембране нервный импульс.
Нервный импульс — это движущаяся волна изменений в состоянии мембраны. Волна включает в себя три вида изменений: структурные (открытие и закрытие мембранных ионных каналов); химические (трансмембранные потоки ионов); электрические (изменения электрического потенциала мембраны) Электрических изменений тоже три: деполяризация, позитивная поляризация (=инверсия, =овершут) и реполяризация. Электрические изменения (проявления) в состоянии мембраны — это потенциал действия. © 2012-2021 Сазонов В.Ф. © 2012-2021 kineziolog.su
Можно сказать короче:
«Нервный импульс — это волна изменений, движущаяся по мембране нейрона». © 2012-2021 Сазонов В.Ф. © 2012-2021 kineziolog.su
Но в физиологической литературе в качестве синонима для нервного импульса принято использовать также и термин «потенциал действия». Хотя потенциал действия — это только электрический компонент нервного импульса.
Потенциал действия – это резкое скачкообразное изменение мембранного потенциала с отрицательного на положительный и обратно.
Сравним два понятия: «нервный импульс» и «потенциал действия».
Нервный импульс — это сложный структурно-электро-химический процесс, распространяющийся по мембране нейрона в виде бегущей волны изменений.
Потенциал действия — это только электрический компонент нервного импульса, характеризующий изменения электрического потенциала на локальном участке мембраны во время прохождения через него нервного импульса. Потенциал мембраны при этом изменяется от -70 до +30 мВ и обратно до -70 мВ — это и есть потенциал действия. (Кликните на изображение слева, чтобы увидеть анимацию.)
Сравните два приведённых выше рисунка (покликайте по ним) и, как говорится, почувствуйте разницу!
Где рождаются нервные импульсы?
Как ни странно, не все студенты, изучившие физиологию возбуждения, могут ответить на этот вопрос. ((
Хотя ответ не сложен. Нервные импульсы рождаются на нейронах всего в нескольких местах:
1) аксонный холмик (это переход тела нейрона в аксон),
2) рецепторное окончание дендрита,
3) первый перехват Ранвье на дендрите (триггерная зона дендрита),
4) постсинаптическая мембрана возбуждающего синапса.
Места возникновения нервных импульсов:
1. Аксонный холмик — главный породитель нервных импульсов
Аксонный холмик — это самое начало аксона, там где он начинается на теле нейрона. Именно аксонный холмик является главным породителем (генератором) нервных импульсов на нейроне. Во всех остальных местах вероятность рождения нервного импульса намного меньше. Дело в том, что у мембраны аксонного холмика повышена чувствительность к возбуждению и понижен критический уровень деполяризации (КУД) по сравнению с остальными участками мембраны. Поэтому, когда на мембране нейрона начинают суммироваться многочисленные возбуждающие постсинаптические потенциалы (ВПСП), которые возникают в самых разных местах на постсинаптических мембранах всех его синаптических контактов, то раньше всего КУД достигается именно на аксонном холмике. Там-то эта сверхпороговая для холмика деполяризация и открывает потенциал-чувствительные натриевые каналы, в которые входит поток ионов натрия, порождающий потенциал действия и нервный импульс.
Итак, аксонный холмик является интегративной зоной на мембране, он интегрирует все возникающие на нейроне локальные потенциалы (возбуждающие и тормозные) — и первый срабатывает на достижение КУД, порождая нервный импульс.
Важно также учесть следующий факт. От аксонного холмика нервный импульс разбегается по всей мембране своего нейрона: как по аксону к пресинаптическоим окончаниям, так и по дендритам к постсинаптическим «начинаниям». Все локальные потенциалы при этом снимаются с мембраны нейрона и со всех его синапсов, т.к. они «перебиваются» потенциалом действия от пробегающего по всей мембране нервного импульса.
2. Рецепторное окончание чувствительного (афферентного) нейрона
Если нейрон имеет рецепторное окончание, то на него может воздействовать адекватный раздражитель и порождать на этом окончании сначала рецепторный потенциал, затем генераторный потенциал, а потом и нервный импульс. Когда генераторный потенциал достигает КУД, то на этом окончании открываются потенциал-зависимые натриевые ионные каналы и рождается потенциал действия и нервный импульс. Нервный импульс бежит по дендриту к телу нейрона, а затем по его аксону к пресинаптическим окончаниям для передачи возбуждения на следующий нейрон. Так работают, к примеру, болевые рецепторы (ноцицепторы), являющиеся дендритными окончаниями болевых нейронов. Нервные импульсы в болевых нейронах вознимают именно на рецепторных окончаниях дендритов.
3. Первый перехват Ранвье на дендрите (триггерная зона дендрита)
Локальные возбуждающие постсинаптические потенциалы (ВПСП) на окончаниях дендрита, которые формируются в ответ на возбуждения, приходящие к дендриту через синапсы, суммируются на первом перехвате Ранвье этого дендрита, если он, конечно, миелинизирован. Там находится участок мембраны с повышенной чувствительностью к возбуждению (пониженным порогом), поэтому именно в этом участке легче всего преодолевается критический уровень деполяризации (КУД), после чего открываются потенциал-управляемые ионные каналы для натрия — и возникает потенциал действия (нервный импульс).
4. Постсинаптическая мембрана возбуждающего синапса
В редких случаях ВПСП на возбуждающем синапсе может быть настолько силён, что прямо там же достигает КУД и порождает нервный импульс. Но чаще это бывает возможно только в результате суммации нескольких ВПСП: или с нескольких соседних синапсов, сработавших одновременно (пространственная суммация), или за счёт того, что на данный синапс пришло несколько импульсов подряд (временная суммация).
Видео: Проведение нервного импульса по нервному волокну
Потенциал действия как нервный импульс
Ниже размещён материал, взятый из учебно-методического пособия автора данного сайта, на который вполне можно ссылаться в своём списке литературы:
Сазонов В.Ф. Понятие и виды торможения в физиологии центральной нервной системы: Учебно-методическое пособие. Ч. 1. Рязань: РГПУ, 2004. 80 с.
Все процессы мембранных изменений, происходящих в ходе распространяющегося возбуждения, достаточно хорошо изучены и описаны в научной и учебной литературе. Но не всегда это описание легко понять, поскольку в данном процессе задействовано слишком много компонентов (с точки зрения обычного студента, а не вундеркинда, конечно).
Для облегчения понимания мы предлагаем рассматривать единый электрохимический процесс распространяющегося динамичного возбуждения с трёх сторон, на трёх уровнях:
- Электрические явления – развитие потенциала действия.
- Химические явления – движение ионных потоков.
- Структурные явления – поведение ионных каналов.
Три стороны процесса
распространяющегося возбуждения
1. Потенциал действия (ПД)
Потенциал действия – это скачкообразное изменение постоянного мембранного потенциала с отрицательной поляризации на положительную и обратно.
Обычно мембранный потенциал в нейронах ЦНС изменяется от –70 мВ до +30 мВ, а затем вновь возвращается к исходному состоянию, т.е. к –70 мВ. Как видим, понятие потенциала действия характеризуется через электрические явления на мембране.
На электрическом уровне
изменения начинаются как смена поляризованного состояния мембраны на деполяризацию, что означает уменьшение электроотрицательности на внутренней стороне мембраны. Сначала деполяризация иёет в виде локального возбуждающего потенциала. Вплоть до критического уровня деполяризации (примерно –50 мВ) это относительно простое линейное уменьшение электроотрицательности, пропорциональное силе воздействующего раздражителя. А вот потом начинается более крутая
самоусиливающаяся
деполяризация, она развивается не с постоянной скоростью, а
с ускорением
. Говоря образно, деполяризация так разгоняется, что в разгону перескакивает через нулевую отметку, не заметив этого, и даже переходит в положительную поляризацию. После достижения пика (обычно +30 мВ) начинается обратный процесс –
реполяризация
, т.е. восстановление отрицательной поляризации мембраны.
Кратко опишем электрические явления во время течения потенциала действия:
Восходящая ветвь графика:
- Потенциал покоя – исходное обычное поляризованное электроотрицательное состояние мембраны (–70 мВ).
- Нарастающий локальный потенциал – пропорциональная раздражителю деполяризация в интервале от –70 мВ до –50 мВ.
- Критический уровень деполяризации (–50 мВ) запускает резкое ускорение деполяризации за счёт самораскрытия нового вида натриевых каналов (потенциал-управляемых), с этой точки начинается спайк – высокоамплитудная часть потенциала действия.
- Самоусиливающаяся круто нарастающая деполяризация за счёт автоматического самораскрытия потенциал-управляемых натриевых каналов.
- Переход нулевой отметки (0 мВ) – смена полярности мембраны. Но новых процессов это событие не вызывает.
- «Овершут» – положительная поляризация (=инверсия, или =реверсия потенциала мембраны). Электроотрицательность переходит в электроположительность.
- Пик (+30 мВ) – вершина процесса изменения полярности мембраны, вершина потенциала действия. Открытые потенциал-управляемые натриевые каналы к этому моменту самостоятельно автоматически закрываются изнутри специальными белковыми «пробками», и поступление положительно заряженных ионов натрия в клетку прекращается. Поэтому прекращается нарастание потенциала действия.
Нисходящая ветвь графика:
- Реполяризация – восстановление прежней исходной электроотрицательности мембраны. Потенциал от +30 мВ опускается вниз. Это происходит благодаря утечке ионов калия из клетки через множество открытых калиевых каналов. Эти ионы выходят из клетки под действием химической силы, из-за разности их концентрации внутри и снаружи клетки. Внутри их много, а снаружи мало, вот они и перемещаются туда, где их мало.
- Переход нулевой отметки (0 мВ) – обратная смена полярности мембраны на прежнюю, отрицательную. Но ничего принципиально нового здесь не происходит. Разве что с этого момента начинает действовать электрическая сила, затягивающая положительные ионы внутрь клетки и удерживающая те из них, которые уже находятся в клетке. Ионам калия с этого момента становится всё труднее выходить из клетки.
- Переход критического уровня деполяризации (–50 мВ) – прекращение фазы относительной рефрактерности (невозбудимости) и возврат возбудимости нейрона.
- Следовые процессы (следовая деполяризация или следовая гиперполяризация).
- Восстановление потенциала покоя – возврат мембраны к своему нормальному состоянию: –70 мВ.
Итак, сначала – деполяризация, затем – реполяризация. Сначала – утрата электроотрицательности, затем – восстановление электроотрицательности.
2. Ионные потоки
Образно можно сказать, что заряженные ионы – это и есть создатели электрических потенциалов в нервных клетках. Для многих людей звучит странно утверждение, что вода не проводит электрический ток. Но на самом деле это так. Сама по себе вода является диэлектриком, а не проводником. В воде электрический ток обеспечивают не электроны, как в металлических проводах, а заряженные ионы: положительные катионы и отрицательные анионы. В живых клетках основную «электрическую работу» выполняют катионы, так как они более подвижны. Электрические токи в клетках – это потоки ионов.
Итак, важно осознать, что все электрические токи, которые идут через мембрану, являются ионными потоками
. Привычного нам из физики тока в виде потока электронов в клетках, как в водных системах, просто нет. Ссылки на потоки электронов будут ошибкой.
На химическом уровне
мы, описывая распространяющееся возбуждение, должны рассмотреть, как изменяются характеристики ионных потоков, идущих через мембрану. Главное в этом процессе то, что при деполяризации резко усиливается поток ионов натрия внутрь клетки, а затем он внезапно прекращается на спайке потенциала действия. Входящий поток натрия как раз и вызывает деполяризацию, так как ионы натрия приносят с собой положительные заряды в клетку (чем и снижают электроотрицательность). Затем, после спайка, значительно нарастает выходящий наружу поток ионов калия, что вызывает реполяризацию. Ведь калий, как мы неоднократно говорили, выносит с собой из клетки положительные заряды. Отрицательные заряды остаются внутри клетки в большинстве, и за счет этого усиливается электроотрицательность. Это и есть восстановление поляризации за счет выходящего потока ионов калия. Заметим, что выходящий поток ионов калия возникает практически одновременно с появлением натриевого потока, но нарастает медленно и длится в 10 раз дольше. Несмотря на продолжительность калиевого потока самих ионов расходуется немного – всего одна миллионная доля от запаса калия в клетке (0,000001 часть).
Подведем итоги. Восходящая ветвь графика потенциала действия образуется за счет входа в клетку ионов натрия, а нисходящая – за счет выхода из клетки ионов калия.
3. Ионные каналы
Все три стороны процесса возбуждения – электрическая, химическая и структурная – необходимы для понимания его сущности. Но все-таки все начинается с работы ионных каналов. Именно состояние ионных каналов предопределяет поведение ионов, а поведение ионов в свою очередь сопровождается электрическими явлениями. Начинают процесс возбуждения натриевые каналы
.
На молекулярно-структурном уровне
происходит открытие мембранных натриевых каналов. Сначала этот процесс идет пропорционально силе внешнего воздействия, а затем становится просто «неудержимым» и массовым. Открытие каналов обеспечивает вход натрия в клетку и вызывает деполяризацию. Затем, примерно через 2-5 миллисекунд, происходит их
автоматическое закрытие
. Это закрытие каналов резко обрывает движение ионов натрия внутрь клетки, и, следовательно, обрывает нарастание электрического потенциала. Рост потенциала прекращается, и на графике мы видим спайк. Это вершина кривой на графике, дальше процесс пойдет уже в обратном направлении. Конечно, очень интересно разобраться в том, что натриевые каналы имеют двое ворот, и открываются они активационными воротами, а закрываются инактивационными, но это следует обсуждать ранее, в теме «Возбуждение». Мы на этом останавливаться не будем.
Параллельно в открытием натриевых каналов с небольшим отставанием во времени идет нарастающее открытие калиевых каналов. Они медлительные по сравнению с натриевыми. Открытие дополнительных калиевых каналов усиливает выход положительных ионов калия из клетки. Выход калия противодействует «натриевой» деполяризации и вызывает восстановление полярности (восстановление электроотрицательности). Но натриевые каналы опережают калиевые, они срабатывают примерно в 10 раз быстрее. Поэтому входящий поток положительных ионов натрия в клетку опережает компенсирующий выход ионов калия. И поэтому деполяризация развивается опережающими темпами по сравнению с противодействующей ей поляризацией, вызванной утечкой ионов калия. Вот почему, пока натриевые каналы не закроются, восстановление поляризации не начнется.
Пожар как метафора распространяющегося возбуждения
Для того чтобы перейти к пониманию смысла динамичного
процесса возбуждения, т.е. к пониманию его распространения вдоль мембраны, надо представить себе, что описанные нами выше процессы захватывают сначала ближайшие, а затем все новые, все более и более отдаленные участки мембраны, пока не пробегут по всей мембране полностью. Если вы видели «живую волну», которую устраивают болельщики на стадионе за счет вставания и приседания, то вам легко будет представить себе мембранную волну возбуждения, которая образуется за счет последовательного протекания в соседних участках трансмембранных ионных токов.
Когда мы искали образный пример, аналогию или метафору, которая может наглядно передать смысл распространяющегося возбуждения, то остановились на образе пожара. Действительно, распространяющееся возбуждение похоже на лесной пожар, когда горящие деревья остаются на месте, а фронт огня распространяется и уходит все дальше и дальше во все стороны от очага возгорания.
Как же в этой метафоре будет выглядеть явление торможения?
Ответ очевиден – торможение будет выглядеть как тушение пожара, как уменьшение горения и затухание огня. Но если огонь распространяется сам по себе, то тушение требует усилий. Из потушенного участка процесс тушения сам по себе не пойдет во все стороны.
Существует три варианта борьбы с пожаром: (1) либо надо ждать, когда все сгорит и огонь истощит все горючие запасы, (2) либо надо поливать водой горящие участки, чтобы они погасли, (3) либо надо поливать заранее ближайшие нетронутые огнем участки, чтобы они не загорелись.
Можно ли «погасить» волну распространяющегося возбуждения?
Вряд ли нервная клетка способна «погасить» этот начавшийся «пожар» возбуждения. Поэтому первый способ подходит только для искусственного вмешательства в работу нейронов (например, в лечебных целях). Но вот «залить водичкой» некоторые участки и поставить блок распространению возбуждения, оказывается, вполне возможно.
© Сазонов В.Ф. Понятие и виды торможения в физиологии центральной нервной системы: Учебно-методическое пособие. Ч. 1. Рязань: РГПУ, 2004. 80 с.
АВТОВОЛНЫ В АКТИВНО-ВОЗБУДИМЫХ СРЕДАХ (АВС)
При распространении волны в активно-возбудимых средах не происходит переноса энергии. Энергия не переносится, а освобождается, когда до участка АВС доходит возбуждение. Можно провести аналогию с серией взрывов зарядов, заложенных на некотором расстоянии друг от друга (например, при тушении лесных пожаров, строительстве, мелиоративных работах), когда взрыв одного заряда вызывает взрыв рядом расположенного и так далее. Лесной пожар также является примером распространения волны в активно- возбудимой среде. Пламя распространяется по области с распределенными запасами энергии — деревья, валежник, сухой мох.
Основные свойства волн, распространяющихся в активно-возбудимых средах (АВС)
Волна возбуждения распространяется в АВС без затухания; прохождение волны возбуждения связано с рефрактерностью — невозбудимостью среды в течение некоторого промежутка времени (периода рефрактерности).
Источник: Антонов В.Ф., 1996.
Смотрите также: Потенциал действия мотонейрона
Видео: Потенциал действия (Action potential)
© 2012-2019 Сазонов В.Ф. © 2016-2019 kineziolog.su.
Распространение потенциала действия между клетками
В химическом синапсе после того, как волна потенциала действия доходит нервного окончания, она вызывает высвобождение нейротрансмиттеров из пресинаптических пузырьков в синаптическую щель. Молекулы медиатора, высвобождаемых с пресинапса, связываются с рецепторами на постсинаптической мембране, в результате чего в рецепторных макромолекулах открываются ионные каналы. Ионы, начинают поступать внутрь постсинаптической клетки через открытые каналы, изменяют заряд ее мембраны, что приводит к частичной деполяризации мембраны и, как следствие, провоцирование генерации постсинаптической клетки потенциала действия.
В электрическом синапсе отсутствует «посредник» передачи в виде нейромедиатора. Зато клетки соединены между собой с помощью специфических протеиновых тоннелей — конексонив, поэтому ионные токи, с пресинаптической клетки могут стимулировать постсинаптическую клетку, вызывая зарождения в ней потенциала действия. Благодаря такому строению, потенциал действия может распространяться в обе стороны и значительно быстрее, чем через химический синапс.
- Схема процесса передачи нервного сигнала в химическом синапсе
- Схема строения электрического синапса
Потенциал действия в различных типах клеток
Потенциал действия в мышечных тканях
Потенциал действия в скелетных мышечных клетках аналогичный потенциала действия в нейронах. Потенциал покоя в них как правило -90мВ, что меньше, чем потенциал покоя типовых нейронов. Потенциал действия мышечных клеток длится примерно 2-4 мс, абсолютный рефрактерный период составляет примерно 1-3 мс, а скорость проводимости вдоль мышц примерно 5 м / с.
Потенциал действия в сердечных тканях
Потенциал действия клеток рабочего миокарда состоит из фазы быстрого деполяризации, начальной быстрой реполяризации, которая переходит в фазу медленной реполяризации (фаза плато), и фазы быстрой конечной реполяризации. Фаза быстрой деполяризации обусловлена резким повышением проницаемости мембраны для ионов натрия, вызывает быстрый входящий натриевый ток, при достижении мембранного потенциала 30-40 мВ инактивируется и в дальнейшем главную роль играют кальциевый ионный ток. Деполяризация мембраны вызывает активацию кальциевых каналов, в результате чего возникает дополнительный Деполяризующий входящий кальциевый ток.
Потенциал действия в сердечной ткани играет важную роль в координации сокращений сердца.
Потенциал действия, его фазы, их происхождение.
Потенциал действия — это быстрое колебание мембранного потенциала возникающее при возбуждении мембраны.
Фазы: 1) медленная деполяризация (так же локальный ответ) — возникает вследствие увеличение проницаемости мембраны для ионов натрия. Под пороговый стимул недостаточен, чтобы вызвать быструю деполяризацию сразу. Длительность фазы зависит от силы раздражителя.
2) быстрая деполяризация — характеризуется быстрым уменьшением мембранного потенциала и даже перезарядкой мембраны (овершут): внутренняя ее часть на некоторое время становится заряженной положительно, а внешняя отрицательно. Это происходит вследствие лавинообразно по ступающего натрия внутрь клетки. В отличие от локального ответа скорость и величина деполяризации не зависит от силы раздражителя. Продолжительность фазы деполяризации в нервном волокне лягушки составляет около 0.2 — 0.5 мс.
3) реполяризация (продолжительность 0.5-0.8 мс) — мембранный потенциал постепенно восстанавливается и достигает 75 — 85% потенциала покоя.
2 и 3 фазы называются пиком потенциала действия.
4) следовая деполяризация — является продолжением фазы реполяризации и характеризуется более медленным (по сравнению с фазой реполяризации) восстановлением потенциала покоя
5) следовая гиперполяризация — представляет собой временное увеличение мембранного потенциала выше исходного уровня.
4 и 5 фазу называют следовыми явлениями
Соотношение фаз возбудимости с фазами потенциала действия. Рефрактерность и ее причины.
Уровень возбудимости клетки зависит от фазы ПД. В фазу локального ответа возбудимость возрастает. Это фазу возбудимости называют латентным дополнением. В фазу деполяризации ПД, когда открываются все натриевые каналы и ноны натрия лавинообразно устремляются в клетку, никакой даже сверхсильный раздражитель не может стимулировать этот процесс. Поэтому фазе деполяризации соответствует фаза полной не возбудимости или абсолютной рефрактерности, т.е. фазе реполяризации все большая часть натриевых каналов закрывается. Однако они могут вновь открываться при действии сверхпорогового раздражителя. Т.е. возбудимость начинает вновь повышаться. Этому соответствует фаза относительной не возбудимости или относительной рефрактерности. Во время следовой деполяризации МП находится у критического уровня, поэтому даже допороговые стимулы могут вызвать возбуждение клетки. Следовательно, в этот момент ее возбудимость повышена. Эта фаза называется фазой экзальтации или супернормальной возбудимости.
В момент следовой гиперполяризации МП выше исходного уровня, т.е. дальше КУД и ее возбудимость снижена. Она наводится в фазе субнормальной возбудимости. Рис. Следует отметить, что явление .аккомодации также связано с изменением проводимости ионных каналов. Если деполяризуюший ток нарастает медленно, то это приводит к частичной инактивации натриевых, и активации калиевых каналов. Поэтому развития ПД не происходит.
Следовые явления, их происхождение.
Следовые явления связаны с восстановительными процессами,которые медленно развиваются в нервных и мышечных волокнах после возбуждения. Различают два вида следовых явлений:
1)Следовой отрицательный потенциал или следовая деполяризация мембраны. Возникновение фазы следовой деполяризации объясняется тем, что небольшая часть медленных натриевых каналов остается открытой. Следовая деполяризация хорошо выражена в мякотных нервных волокнах.
2)Следовой положительный потенциал или следовая гиперполяризация мембраны. Следовая гиперполяризация связана с повышенной, после ПД, калиевой проводимостью мембраны и тем, что более активно работает натрий-калиезый насос, выносящий вошедшие в клетку во время ПД ионы натрия. Следовая гиперполяризация хорошо выражена в безмякотных нервных волокнах.
Местное и распространяющееся возбуждение. Локальный ответ
Возбуждение может быть 2-х видов:
-местное (локальный ответ);
-распространяющееся (импульсное).
Местное возбуждение — наиболее древний вид (низшие формы организмов и низковозбудимые ткани — например, соединительная ткань). Местное возбуждение возникает и в высокоорганизованных тканях под действием подпорогового раздражителя или как компонент потенциала действия. При местном возбуждении нет видимой ответной реакции.
Особенности местного возбуждения:
-нет латентного (скрытого) периода — возникает сразу же при действии раздражителя;
-нет порога раздражения;
-местное возбуждение градуально — изменение заряда клеточной мембраны пропорционально силе подпорогового раздражителя;
-нет рефрактерного периода, наоборот характерно небольшое повышение возбудимости;
-распространяется с декрементом (затуханием).
Импульсное (распространяющееся) возбуждение — присуще высокоорганизменным тканям, возникает под действием порогового и сверхпорогового раздражителей.
Особенности импульсного возбуждения: -имеет латентный период — между моментом нанесения раздражения и видимой ответной реакцией проходит некоторое время; -имеет порог раздражения; -не градуально — изменение заряда клеточной мембраны не зависит от силы раздражителя; -наличие рефрактерного периода; -импульсное возбуждение не затухает.
Локальный ответ (ЛО) – активная реакция клетки на электрический раздражитель, однако состояние ионных каналов и транспорт ионов при этом изменяется незначительно. ЛО не проявляется заметной физиологической реакцией клетки. ЛО называют местным возбуждением, так как это возбуждение не распространяется по мембранам возбудимых клеток.
Молекулярные механизмы возникновения потенциала действия
Активные свойства мембраны, обеспечивающие возникновение потенциала действия, основанные главным образом на поведении потенциал-зависимых натриевых (Na +) и калиевых (K +) каналов. Начальная фаза ПД формируется входным натриевым током, позже открываются калиевые каналы и выходной K + -ток возвращает потенциал мембраны к исходному уровню. Начальное концентрацию ионов затем восстанавливает натрий-калиевый насос.
По ходу ПД каналы переходят из состояния в состояние: в Na + -каналов основных состояния трех — закрытый, открытый и инактивированный (в реальности все сложнее, но этих трех состояний достаточно для описания), в K + каналов два — закрытый и открытый.
Поведение каналов, участвующих в формировании ПД, описывается через проводимость и рассчитывается через коэффициенты переноса (трансфера).
Коэффициенты переноса были выведены Алан Ллойд Ходжкин и Эндрю Хаксли.
Проводимость для калия G K на единицу площади
| , |
| где: |
| an — Коэффициент трансфера из закрытого в открытое состояние для K + каналов [1 / s]; |
| bn — Коэффициент трансфера из открытого в закрытое состояние для K + каналов [1 / s]; |
| n — Фракция К + каналов в открытом состоянии; |
| (1 — n) — Фракция К + каналов в закрытом состоянии |
Проводимость для натрия G Na на единицу площади
рассчитать сложнее, поскольку, как уже было упомянуто, в потенциал-зависимых Na + каналов, кроме закрытого / открытого состояний, переход между которыми параметром, еще инактивированный / никак инактивированный состояния, переход между которыми описывается через параметр
| , | , |
| где: | где: |
| am — Коэффициент трансфера из закрытого в открытое состояние для Na + каналов [1 / s]; | ah — Коэффициент трансфера из инактивированного в не-инактивированный состояние для Na + каналов [1 / s]; |
| bm — Коэффициент трансфера из открытого в закрытое состояние для Na + каналов [1 / s]; | bh — Коэффициент трансфера из не-инактивированного в инактивированный состояние для Na + каналов [1 / s]; |
| m — Фракция Na + каналов в открытом состоянии; | h — Фракция Na + каналов в не-инактивированном состоянии; |
| (1 — m) — Фракция Na + каналов в закрытом состоянии | (1 — h) — Фракция Na + каналов в инактивированном состоянии. |
История
Основные положения мембранной теории возбуждения сформулированы немецким нейрофизиологом Ю.Бернштейном
В 1902 году Юлиус Бернштейн выдвинул гипотезу, согласно которой клеточная мембрана пропускает внутрь клетки ионы К +, и они накапливаются в цитоплазме. Расчет величины потенциала покоя по уравнению Нернста для калиевого электрода удовлетворительно совпал с измеренным потенциалом между саркоплазме мышцы и окружающей средой, который составил около — 70 мВ. Согласно теории Ю.Бернштейна, при возбуждении клетки ее мембрана повреждается, и ионы К + выходят из клетки по концентрационном градиента до тех пор, пока потенциал мембраны не становится равным нулю. Затем мембрана восстанавливает свою целостность, и потенциал возвращается к уровню потенциала покоя.
Эту модель развили в своей работе 1952 Алан Ллойд Ходжкин и Эндрю Хаксли в которой описали электрические механизмы, обусловливающие генерацию и передачу нервного сигнала в гигантском аксоне кальмара. За это авторы модели получили Нобелевскую премию в области физиологии и медицины за 1963 год. Модель получила название модель Ходжкина-Хаксли
В 2005 году Томасом Геймбургом и Анрю Д. Джексоном предложена солитонном модель, основанная на предположении, что сигнал по нейронам распространяется в виде солитонов — устойчивых волн, распространяющихся по клеточной мембране.
Влияние некоторых веществ на потенциал действия
Некоторые вещества органического или синтетического происхождения могут блокировать образование или прохождения ПД:
- Батрахотоксин найден у некоторых представителей рода листолазов. Устойчиво и необратимо повышает проницаемость мембран для ионов натрия.
- Понератоксин был найден в муравьях рода Paraponera. Блокирует натриевые каналы.
- Тетродотоксин найден в тканях рыб семейства Скелезубови, из которых готовят японский деликатес Фугу. Блокирует натриевые каналы.
- Механизм действия большинства анестетиков (Прокаин, Лидокаин) базируется на блокировании натриевых каналов и соответственно на блокировании проведении импульсов по чувствительным нервным волокнам.
- 4-Аминопиридин — обратно блокирует калиевые каналы, удлиняет срок потенциала действия. Может использоваться в терапии рассеянного склероза.
- ADWX 1 — обратно блокирует калиевые каналы. В условиях опыта облегчал течение острого рассеянного энцефаломиелита у крыс.