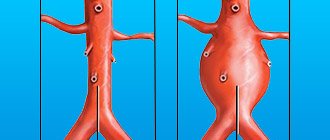
Особенности строения гладких мышц
Мышцы данной группы находятся практически во всех важных внутренних органах, таких, как кишечник, желудок, матка, также они присутствуют в стенках сосудов, коже и глазах. Гладкая мускулатура выполняет непроизвольные движения, подчиняясь лишь автоматическим сигналам нервной системы.
Клетки мышц имеют веретенообразную форму, они укорачиваются вследствие скольжения своих нитей. Скорость этого процесса гораздо медленнее, чем у скелетных мышц, благодаря чему они способны долгое время находиться в напряженном состоянии, не затрачивая для этого много энергии.
Важной особенностью гладких мышц является их способность сохранять форму, измененную растяжением или деформацией, а также высокая пластичность, что немаловажно для работы внутренних органов. Мышцы этой группы характеризуются самым медленным сокращением и расслаблением, которое может продолжаться до нескольких десятков секунд. Также они могут долгое время находиться в состоянии тонуса, практически не утомляясь. Основные функции гладкой мускулатуры: •
- поддержание давления в полых внутренних органах (мочеточник, кишечник, матка);
- сокращаясь, они обеспечивают естественную перистальтику органов и их опорожнение;
- регулируют давление в кровеносных сосудах;
- в органах зрения обеспечивают расширение и сужение зрачка;
- расположенные на кожных покровах, они способствуют выделению подкожного жира.
Продолжительное растяжение гладких мышц приводит к их напряжению, что, в свою очередь, играет важную роль в продвижении содержимого органов желудочно-кишечного тракта и мочеточников. Гладкие мышцы стенок кровеносных сосудов, сокращаясь, оказывают влияние на кровоснабжение организма и артериальное давление.
Физиологические свойства и особенности сердечной мышцы
По морфологическим и функциональным признакам в сердце выделяют два типа мышечных волокон:
- волокна рабочего миокарда предсердий и желудочков;
- волокна водителей ритма (пейсмеккеров) и проводящей системы.
Рабочий миокард электрофизиологически отличается от скелет- ной мышцы. Потенциал покоя типических кардиомиоцитов калиевой природы. Его величина составляет около -90 мв, уровень критической деполяризации – 50 мв.
Потенциал действия имеет амплитуду 120 мв и длительность 200- 400 мс. В нем выделяют фазы: 0 – быстрой деполяризации, 1- начальной быстрой реполяризации, 2- медленной реполяризации, 3- быстрой реполяризации, 4- фазу покоя. Увеличение частоты сокращения сердца приводит к снижению продолжительности 3 и 4 фаз при неизменной длительности 0, 1 и 2 (рис. 54).
Фаза быстрой деполяризации связана с увеличением тока натрия и кальция через натриевые и быстрые кальциевые каналы. Селективность последних относительная, т.к. через них могут проходить катионы натрия при дефиците кальция. По достижению потенциала в -30 – -40 мв натриевые каналы инактивируются. Плато потенциала действия связано с активацией кальциевых каналов. Высокий кальциевый ток уменьшает калиевую проводимость мембраны, что поддерживает ее деполяризованное состояние.
В конце 2 фазы кальциевая проводимость мембраны уменьшается и увеличивается калиевая, ток которого и определяет процесс быстрой реполяризации (3 фаза). В состоянии покоя деполяризующие токи кальция и натрия уравновешены с реполяризующим током калия. Потенциал действия типических кардиомиоцитов вызванный (возникает в ответ на раздражение).
При возникновении и развитии потенциала действия возникают изменения возбудимости. Быстрая деполяризация сопровождается абсолютной рефрактерностью ( 0,27 с ). Причина – инактивация натриевых каналов. Они начинают восстанавливаться при достижении в процессе реполяризации заряда в -60 – -70 мв. Наступает фаза относительной рефрактерности ( 0,03 с ).
Вслед за фазой относительной рефрактерности возможны кратковременные фазы экзальтации и пониженной фозбудимости.
Функционально значение большого периода абсолютной рефрактерности состоит в том, что предотвращается возможность циркуляции возбуждения по сердцу (рефрактерный период больше, чем время про- ведения возбуждения по сердцу) и его тетаническому сокращению при ритмическом раздражении.
В нормальном сердце возбуждения и сокращения рабочего миокарда вызываются потенциалами действия, которые приходят по проводящей системе сердца от водителей ритма.
Атипические кардиомиоциты расположены в узлах и пучках проводящей системы сердца. Они способны к спонтанному возбуждению. В период диастолы сердца в них возникают волны деполяризации – медленная диастолическая деполяризация, которая лежит в основе автоматии сердца.
Электрофизиологически пейсмеккерные клетки характеризуются низким потенциалом покоя (-50 – -60 мв) и низким уровнем критической деполяризации (-40 -50 мв). В покое натриевая проводимость мембраны больше, чем в скелетной мышце и в типических кардиомиоцитах ( Р + : P +: P _= 0,48 : 1 : 0,02 ). Увеличенная натриевая проводимость мембраны способствует их высокой возбудимости.
В отличие от типических кардиомиоцитов, атипические способны к самовозбуждению, в основе которого лежит их способность к медленной диастолической деполяризации. Потенциал действия атипических кардиомиоцитов натрий-кальциевой природы. Его амплитуда составляет 60-70 мв.
Таким образом, в настоящее время общепризнанной является миогенная теория автоматии, согласно которой возбуждение и сокращение сердца связано с возникновением потенциалов действия в атипических кардиомиоцитах и их распространении по проводящей системе на рабочий миокард.
Атипические кардиомиоциты обладают разной способностью к самовозбуждению. В пределах водителей ритма сердца (например в синатриальном узле) находятся как ведущие пейсмеккеры (определяют частоту возбуждения сердца), так и латентные (при работающих ведущих пейсмеккерах выполняют подчиненную функцию). Латентные пейсмеккеры включаются в работу при нарушении функций ведущих.
Разная способность атипических кардиомиоцитов к спонтанной генерации потенциалов действия отражается в разной способности водителей ритма сердца к самовозбуждению. Так, частота потенциалов действия, генерируемых синатриальным узлом, составляет 60-80 в ми- нуту, атриовентрикулярным – 40-60, волокнами пучка Гиса 30-40.
Поэтому в нормально работающем сердце возбуждение возникает в синатриальном узле ( этот узел называется нормально расположенным – нормотопным, ритм сердца – синусовым) и навязывается другим участкам проводящей системы, а при его блокаде может возникать в атриовентрикулярном ( этот водитель ритма, также как и пучек Гиса, называют гетеротопным ).
Проведение возбуждения по сердцу осуществляется с разной скоростью. От синатриального узла к атриовентрикулярному возбуждение распространяется по пучкам Бахмана, Венкебаха и Тореля со скоростью 1 м/с, в атриовентрикулярном узле она падает до 0,02-0,04 м/с (атриовентрикулярная задержка ). По пучку Гиса возбуждение распространяется со скоростью 4-2 м/с, и по волокнам Пуркинье – около 2 м/с (рис. 55).
Сопряжение возбуждения и сокращения в сердце.
Ключевым событием начала сокращения служит вход кальция в клетку. Входящий кальций увеличивает продолжительность потенциала действия и пополняет запасы внутриклеточного кальция.
Таким образом, потенциал действия не только вызывает процесс сокращения (как в скелетной мышце), но и влияет на силу сокращения, пополняя запасы кальция.
Чем больше длительность потенциала действия, тем больше амплитуда сокращения. Целый ряд регуляторов деятельности сердца оказывает влияние посредством изменения входящего тока кальция.
Физиологические свойства миокарда:
- возбудимость;
- сократимость;
- проводимость.
Физиологические особенности миокрада:
- большая продолжительность потенциала действия;
- потенциал действия натрий-кальциевой природы;
- большой период рефрактерности;
- меньшая возбудимость;
- меньшая сократимость;
- меньшая проводимость;
- большой латентный период;
- способность к автоматии.
В отличие от скелетной мышцы, кардиомиоциты связаны вставочными дисками (электрическими синапсами), объединяющими их в функциональный синцитий.
Возбуждение, возникающее в одном участке сердца, распространяется на все остальные – сердце подчиняется правилу ―всё или ничего.
Проводящая система сердца
К проводящей системе сердца относят следующие отделы:
1. Синусно-предсердный (синоатриальный узел):
- располагается под правым ушком у места впадения верхней полой вены в правое предсердие,
- находится под эпикардом,
- площадь 20*2 мм^2,
- состоит из 40 тыс. клеток,
- обильно снабжен капиллярами и нервами.
2. Межпредсердные и межузловые проводящие пути — передают возбуждение по предсердиям.
Их выделяют 3:
- передний (пучок Бахмана),
- средний (Веннебаха),
- задний (Торела).
3. Предсердно-желудочковый узел (атрио-вентрикулярный):
- располагается в нижней части межпредсердной перегородки,
- под эндокардом правого предсердия,
- иннервируется волокнами блуждающего и симпатического нервов.
4. Пучок Гиса отходит от атрио-вентрикулярного узла:
- длина 8-10 мм,
- идет по межжелудочковой перегородке,
- на ее вершине раздваивается на правую и левую ножки.
5. Волокна Пуркинье:
- сеть атипичных волокон в стенках обоих желудочков,
- с них передается возбуждение на сократительный миокард желудочков.
Проводящая система сердца:
- атипичные кардиомиоциты,
- клетки богаты саркоплазмой,
- поперечная исчерченность в них выражена менее четко,
- мало миофибрилл,
- сохраняет признаки эмбрионального миокарда,
- устойчива к гипоксии,
- энергия образуется за счет активации процессов анаэробного гликолиза.
Во время диастолы в клетках синоатриального узла (водитель ритма I порядка — пейсмейкер):
- уменьшается мембранный потенциал, то есть происходит медленная диастолическая деполяризация (МДД);
- мембранный потенциал (МП) достигает КУД, то есть МП изменяется с 50-60 мВ до 30-40 мВ самопроизвольно — потенциал действия (ПД) или пейсмекерный потенциал, который распространяется по проводящей системе сердца, переходит на миокард.
Особенности пейсмекерных клеток:
- низкий уровень мембранного потенциала (-50 — -60 мВ),
- способность к МДД (снижению МП до КУД самопроизвольно),
- низкая амплитуда ПД (-30 — -50 мВ) без реверсии (в основном).
Причины МДД (связана с особыми свойствами мембраны пейсмейкеров):
- постепенное самопроизвольное увеличение в диастолу проницаемости мембраны для Na и Ca, входящих в клетку;
- уменьшение проницаемости K, выходящую из клетки;
- уменьшение активности Na-K насоса (Na-K-АТФ-азы).
Частота возбуждений в клетках синоатриального узла — 60-80 за 1 мин. Это водитель ритма I порядка.
Способностью к автоматии обладают все нижележащие проводящие системы сердца (атриовентрикулярный узел, пучок Гиса, волокна Пуркинье, атипичные волокна предсердия). Они являются в норме только потенциальными или латентными водителями ритма.
У атриовентрикулярного узла способность к автоматии — 40-50 имп/мин. Это водитель ритма II порядка.
Клетки пучка Гиса — 30-40 имп/мин.
Волокна Пуркинье — около 20 имп/мин.
В. Гаскелл ввел понятие о градиенте автоматии:
Чем дальше расположен очаг автоматии от венозного конца сердца и ближе к артериальному, тем меньшей способностью к автоматии он обладает
Истинным водителем ритма является клетки синоатриального узла.
Физиологические свойства сердечной мышцы.
Кардиомиоциты изолированы друг от друга и контактируют в области вставочных дисков, где соприкасаются мембраны соседних кардиомиоциов.
Коннесксоны- в мембране соседних клеток образуются эти структуры за счет белков конексинов. Коннексон окружают 6 таких белков, внутри коннексона – канал, который позволяет проходит ионам, таким таким образом электрический ток распространяется от одной клетки к другой. “f область имеет сопротивление 1,4 ом на см2(низкое). Возбуждение охватывает кардиомиоциты одновременно. Они функционирую как функциональный сенсициы. Нексусы очень чувствительны к недостатку кислорода, к действию катехоламинов, к стрессовым ситуациям, к физической нагрузке. Это может вызывать нарушение проведения возбуждения в миокарде. В экспериментальных условиях нарушение плотных контактов моно получить при помещении кусочков миокарда в гипертонический раствор сахарозы. Для ритмической деятельности сердца важна проводящая система сердца – эта система состоит из комплекса мышечных клеток, образующих пучки и узлы и клетки проводящей системы отличаются от клеток рабочего миокарда – они бедны миофибриллами, богаты саркоплазмой и содержат высокое содержание гликогена. Эти особенности при световой микроскопии делают их более светлыми с малой поперечной исчерченностью и они были названы атипическими клетками.
В состав проводящей системы входят:
1. Синоатриальный узел(Кейт-Флека)(в парвом предсердии у места впадения верхней полой вены)
2. Атрии-вентрикулярный узел(Ашоф-Тавара)(лежит в правом предсердии на границе предсердие-желудочек – задняя стенка правого предсердия)
Эти два узла связаны внутрипредсердными трактами —
3. Предсердные тракты
— пердний с ветвью Бахмена к левому предсердию
— средний тракт(Венкебаха)
— задний тракт(Тореля)
4. Пучок Гиса(отходит от атриовентрикулярного узла. Проходит через фиброзную ткань и обеспечивает связь миокарда предсердия с миокардом желудочка. Проходит в межжелудочковую перегородку, где разделяется на правую и илевую ножку пучка Гиса)
5. Правая и левая ножки пучка Гиса(они идут вдоль межжелудочковой перегородки. Левая ножка имеет две ветви – переднюю и заднюю. Конечными разветвлениями будут являтся волокна Пуркинье)
6. Волокна Пуркинье
В проводящей системе сердца, которая образована видоизмененными типами мышечных клеток имеются три вида клеток – пейсмекерны(P), переходные, клетки Пуркинье.
1. P-клетки. Находятся в сино-артриальном узле, меньше в атриовентрикулярном ядре. Это самые мелкие клетки, в них мало т – фибрилл и митохондрий, т-сстема отстутствует, l. система развита слабо. Основной функцией этих клеток является генерация потенциала действия за счет врожденного свойства медленной диастолической деполяризации. В них происходит периодическое снижение мембранного потенциала, которое приводит их к самовозбуждению.
2. Переходные клетки осуществляют предачу возбуждения в области атривентрикуярного ядра. Они обнаруживаются между P клетками и клетками Пуркинье. Эти клетки вытянутой формы, у них отсутствует сарко-плазматический ретикулум. Эти клетки облают замедленной скоростью проведения.
3. Клетки Пуркинье широкие и короткие, в них больше миофибрилл, лучше развит саркоплазматический ретикулум, T система отсутствует.
Электрические свойства клеток миокарда. Клетки миокарда, как рабочего, так и проводящей системы обладают мембранным потенциалам покоя и снаружи мембрана кардиомиоцита заряжена «+», а внутри «-». Это обусловлено ионной ассиметрией – внутри клеток в 30 раз больше ионов калия, а снаружи в 20-25 раз больше ионов натрия. Это обеспечивается постоянной работой натриево-калиевым насосом. Измерение мембранного потенциала показывает, что клетки рабочего миокарда имеетпотенциал – 80-90 мВольт. В клетках проводящей системы – 50-70 мВольт. При возбуждении клеток рабочего миокарда возникает потенциал действия(5 фаз) – 0, 1, 2, 3, 4.
0. При возбуждении возникает процесс деполяризации кардиомиоцитов, что связано с открытием натриевых каналов и повышение проницаемости для ионов натрия, которые устремляются внутрь кардиомиоцитов. При снижении мембранного потенциала о 30-40 милиВольт происходить открытие медленных натриево-кальцевых каналов. Через них могут входить натрий и дополнительно кальций. Это обеспечивает процесс деполяризации и овершут(реверсия) 120 мВольт.
1. Начальная фаза реполяризации. Закрытие натриевых каналов и некоторое повышение проницаемости к ионам хлора.
2. Фаза Плато. Процесс деполяризации затормаживается. Связана с усилением выхода кальция внутрь. Он задерживает восстановление заряда на мембране. При возбуждении снижается калиевая проницаемость(в 5 раз). Калий не может выходить из кардиомиоцитов.
3. Когда кальцевые каналы закрываются происходит фаза быстрой реполяризации. За счет восстановления поляризации к ионам калия и мембранный потенциал возвращается к исходному уровню и наступает диастолический потенциал
4. Диастолический потенциал постоянно стабилен
В клетках проводящей системы есть отличительные особенности потенциала.
1. Сниженный мембранный потенциал в диастолический период(50-70мВ)
2. Четвертая фаза не является стабильной и отмечается постепенное снижение мембранного потенциала к пороговому критическому уровню деполяризации и в диастолу постепенно медленно продолжает снижаться достигая критического уровня деполяризации при котором произойдет самовозбуждение П-клеток. В P-клетках происходит усиление проникновения ионов натрия и снижение выхода ионов калия. Повышается проницаемость ионов кальция. Эти сдвиги в ионном составе приводят к тому, что мембранный потенциал в P-клетках снижается до порогового уровня и p-клетка самовозбуждается обеспечивая возникновение потенциала действия. Плохо выражена фаза Плато. Фаза ноль плавно переходи ТВ процесс реполяризации, который восстанавливает диастолический мембранный потенциал, а дальше цикл повторяется вновь и P-клетки переходят в состояние возбуждения. Наибольшой возбудимостью обладают клетки сино-атриального узла. Потенциал в нем особо низок и скорость диастолической деполяризации наиболее высок.. Это будет влиять на частоту возбуждения. P- клетки синусного узла генерируют частоту до 100 ударов в мин. Нервная система(симпатическая система) подавляют действие узла(70 ударов). Симпатическая система может повышать автоматию. Гуморальные факторы- адреналин, норадреналин. Физические факторы – механический фактор – растяжение, стимулируют автоматию, согревание, тоже увеличивает автоматию. Все это применяется в медицине. На этом основано мероприятие прямого и непрямого массажа сердца. Область атриовентрикулярного узла тоже обладает автоматией. Степень автоматии атриовентрикулярного узла выражена значительно меньше и как правило она в 2 раза меньше, чем в синусном узле – 35-40. В проводящей системе желудочков импульсы тоже могут возникать(20-30 в минуту). ПО ходу проводящей системы возникает постипенное снижение уровня автоматии, что получило название градиента автоматии. Синусный узел – центр автоматии первого порядка.
Станеус – ученый. Наложение лигатур на сердце лягушки(3х камерное). У правого предсердия имеется венозныц синус, где лежит аналог синусного узла человека. Станеус накладывал 1ую лигатуру между венозным синусом и предсердием. Когда лигатура затягивалась сердце прекращала свою работу. Вторая лигатура накладывалась Станеусом между предсердиями и желудочком. В этой зоне находится аналог атрии-вентрикулярного узла, но 2ая лигатура имеет задачу не отделения узла, а его механическое возбуждение. Ее накладывают постепенно, возбуждая атриовентрикулярный узел и при этом возникает сокраение сердца. Желудочки получают вновь сокращаться под действием атрии-вентрикулярного узла. С частотой в 2 раза меньше. Если наложить 3ю лигатуру, которая отделяет атривентрикулярный узел возникает остановка сердца. Все это дает нам возможность показать, что синусный узел – водитель ритма, атриовентрикулярный узел обладает меньшей автоматией. В проводящей системе существуе убывающий градиент автоматии.
Физиологические свойства сердечной мышцы.
Возбудимость, проводимость ,сократимость
Под возбудимостью сердечной мышцы понимается ее свойство отвечать на действие раздражителей пороговой или над пороговой силы процессом возбуждения. Возбуждение миокарда можно получить на действие химических, механических, температурных раздражений. Эта способность отвечать на действие разных раздражителей используется при массаже сердца(механическое), введение адреналина, кардиостимуляторы. Особенностью реакции сердца на действие раздражителя, играет то что действует по принципу «Все или ничего». Сердце отвечает максимальным импульсом уже на пороговый раздражитель. Продолжительность сокращения миокарда в желудочкх составляет 0,3с. Это обусловлено длительным потенциалом действия, который тоже длится до 300мс. Возбудимость сердечной мышцы может падать до 0 – абсолютно рефрактерная фаза. Никакие раздражители не могут вызвать повторного возбуждения(0,25-0,27с). Сердечная мышца абсолютно невозбудима. В момент расслабления(диастолы)абсолютная рефрактерная переходит в относительную рефрактерную 0,03-0,05с. В этот момент можно получить повторное раздражение на над пороговые раздражители. Рефрактерный период сердечной мышцы длится и совпадает по времени столько, сколько длится сокращение. Вслед за относительной рефрактерностью имеется небольшой период повышенной возбудимости – возбудимость ставновится выше исходного уровня – супер нормальная возбудимость. В эту фазу сердце особо чувствительно к воздействию других раздражителей(смогут возникать др. раздражители или экстрасистолы- внеочередные систолы). Наличие длительного рефрактерного периода должно оградить сердце от повторных возбуждений. Сердце выполняет насосную функцию. Промежуток между нормальным и внеочередным сокращением укорачивается. Пауза может быть нормальной или удлиненной.Удлиненную паузу называют компенсаторной. Причина экстрасистолов – возникновение других очагов возбуждения – атриовентрикулярный узел, элементы желудочковой части проводящей системы, клетки рабочего миокарда, Это может быть связано с нарушением кровоснабжением, нарушением проведения в сердечной мышцей, но все дополнительные очаги – эктопические очаги возбуждения. В зависимости от локализации – разные экстрасистолы – синусные, предсредные, атриовентрикулярные. Экстрасистолы желудочка сопровождаются удлиненной компенсаторнйо фазой. 3 дополнительное раздражение – причина внеочередного сокращения. Вовремя экстрасистола сердце утрачивает возбудимость. К ним приходит очередной импульс из синусного узла. Пауза нужна для восстановления нормального ритма. Когда в сердце происходит сбой сердце пропускает одно нормальное сокращение и дальше возвращается к нормальному ритму.
Проводимость – способность проводить возбуждение. Скорость проведения возбуждения в разных отделах неодинакова. В миокарде предсердий – 1 м/c и время проведения возбуждения занимает 0,035 с
Скорость проведения возбуждения
Миокард 1 м/c 0,035
A-V узел 0,02 – 0-05 м в с. 0,04 с
Проведение система желудочков – 2-4,2 м в с. 0,32
В сумме от синусного узла до миокарда желудочка – 0,107 с
Миокард желудочка – 0,8-0,9 м в с
Нарушение проведения сердца приводит к развитию блокад – синусной , атривентрикулярной, пучка гиса и его ножек. Синусный узел может выключится.. Включится ли атривентрикулярный узел как водитель ритма? Синусные блокады встречаются редко. Больше в атриовентрикулярных узлах. Удлиение задержки(больше 0,21с) возбуждение доходит до желудочка, хоть и замедленно. Выпадение отдельных возбуждений, которые возникают в синусном узле НАПРИМЕР из 3 доходит только 2 – вторая степень блокады. 3я блокада – предсердия и желудочки работают несогласованно. Блокада ножек и пучка – блокада желудочков. Чаще встречаются блокады ножек пучка Гиса и соответственно желудочек запаздывает за другим.
Сократимость
Кардиомиоциты включают фибриллы, саркомеры. Есть продольные трубочки и Т трубочки наружной мембраны, котоыре входят внутрь на уровне мембраны я. Они широкие. Сократительная фугкция кардиомиоцитов связана с белками миозином и актином. На тонких актиновых белках – система тропонин и тропомиозин. Это не дает головкам миозин сцепляется с головками миозина. Снятие блокировки — ионами кальция. По т трубочкам открываются кальцевые каналы. Повышение кальция в саркоплазме снимает тормозной эффект актина и миозина. Мостики миозина перемещают тонике нити к центру. Миокард подчиняется в сократительной функции 2м законам – все или ничего. Сила сокращения зависит от исходной длины кардиомиоцитов – Франк и Старалинг. Если миоциты предварительно растянуты, то они отвечают большей силой сокращения. Растяжение зависит от наполнения кровью. Чем больше- тем сильней. Этот закон формулируют как – систола есть функция диастолы. Это важный приспособительный механизм. Это синхронизирует работу правого и левого желудочка.