Физиологические свойства сердечной мышцы.
Кардиомиоциты изолированы друг от друга и контактируют в области вставочных дисков, где соприкасаются мембраны соседних кардиомиоциов.
Коннесксоны- в мембране соседних клеток образуются эти структуры за счет белков конексинов. Коннексон окружают 6 таких белков, внутри коннексона – канал, который позволяет проходит ионам, таким таким образом электрический ток распространяется от одной клетки к другой. “f область имеет сопротивление 1,4 ом на см2(низкое). Возбуждение охватывает кардиомиоциты одновременно. Они функционирую как функциональный сенсициы. Нексусы очень чувствительны к недостатку кислорода, к действию катехоламинов, к стрессовым ситуациям, к физической нагрузке. Это может вызывать нарушение проведения возбуждения в миокарде. В экспериментальных условиях нарушение плотных контактов моно получить при помещении кусочков миокарда в гипертонический раствор сахарозы. Для ритмической деятельности сердца важна проводящая система сердца – эта система состоит из комплекса мышечных клеток, образующих пучки и узлы и клетки проводящей системы отличаются от клеток рабочего миокарда – они бедны миофибриллами, богаты саркоплазмой и содержат высокое содержание гликогена. Эти особенности при световой микроскопии делают их более светлыми с малой поперечной исчерченностью и они были названы атипическими клетками.
В состав проводящей системы входят:
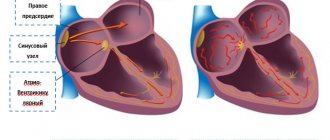
1. Синоатриальный узел(Кейт-Флека)(в парвом предсердии у места впадения верхней полой вены)
2. Атрии-вентрикулярный узел(Ашоф-Тавара)(лежит в правом предсердии на границе предсердие-желудочек – задняя стенка правого предсердия)
Эти два узла связаны внутрипредсердными трактами —
3. Предсердные тракты
— пердний с ветвью Бахмена к левому предсердию
— средний тракт(Венкебаха)
— задний тракт(Тореля)
4. Пучок Гиса(отходит от атриовентрикулярного узла. Проходит через фиброзную ткань и обеспечивает связь миокарда предсердия с миокардом желудочка. Проходит в межжелудочковую перегородку, где разделяется на правую и илевую ножку пучка Гиса)
5. Правая и левая ножки пучка Гиса(они идут вдоль межжелудочковой перегородки. Левая ножка имеет две ветви – переднюю и заднюю. Конечными разветвлениями будут являтся волокна Пуркинье)
6. Волокна Пуркинье
В проводящей системе сердца, которая образована видоизмененными типами мышечных клеток имеются три вида клеток – пейсмекерны(P), переходные, клетки Пуркинье.
1. P-клетки. Находятся в сино-артриальном узле, меньше в атриовентрикулярном ядре. Это самые мелкие клетки, в них мало т – фибрилл и митохондрий, т-сстема отстутствует, l. система развита слабо. Основной функцией этих клеток является генерация потенциала действия за счет врожденного свойства медленной диастолической деполяризации. В них происходит периодическое снижение мембранного потенциала, которое приводит их к самовозбуждению.
2. Переходные клетки осуществляют предачу возбуждения в области атривентрикуярного ядра. Они обнаруживаются между P клетками и клетками Пуркинье. Эти клетки вытянутой формы, у них отсутствует сарко-плазматический ретикулум. Эти клетки облают замедленной скоростью проведения.
3. Клетки Пуркинье широкие и короткие, в них больше миофибрилл, лучше развит саркоплазматический ретикулум, T система отсутствует.
Электрические свойства клеток миокарда. Клетки миокарда, как рабочего, так и проводящей системы обладают мембранным потенциалам покоя и снаружи мембрана кардиомиоцита заряжена «+», а внутри «-». Это обусловлено ионной ассиметрией – внутри клеток в 30 раз больше ионов калия, а снаружи в 20-25 раз больше ионов натрия. Это обеспечивается постоянной работой натриево-калиевым насосом. Измерение мембранного потенциала показывает, что клетки рабочего миокарда имеетпотенциал – 80-90 мВольт. В клетках проводящей системы – 50-70 мВольт. При возбуждении клеток рабочего миокарда возникает потенциал действия(5 фаз) – 0, 1, 2, 3, 4.
0. При возбуждении возникает процесс деполяризации кардиомиоцитов, что связано с открытием натриевых каналов и повышение проницаемости для ионов натрия, которые устремляются внутрь кардиомиоцитов. При снижении мембранного потенциала о 30-40 милиВольт происходить открытие медленных натриево-кальцевых каналов. Через них могут входить натрий и дополнительно кальций. Это обеспечивает процесс деполяризации и овершут(реверсия) 120 мВольт.
1. Начальная фаза реполяризации. Закрытие натриевых каналов и некоторое повышение проницаемости к ионам хлора.
2. Фаза Плато. Процесс деполяризации затормаживается. Связана с усилением выхода кальция внутрь. Он задерживает восстановление заряда на мембране. При возбуждении снижается калиевая проницаемость(в 5 раз). Калий не может выходить из кардиомиоцитов.
3. Когда кальцевые каналы закрываются происходит фаза быстрой реполяризации. За счет восстановления поляризации к ионам калия и мембранный потенциал возвращается к исходному уровню и наступает диастолический потенциал
4. Диастолический потенциал постоянно стабилен
В клетках проводящей системы есть отличительные особенности потенциала.
1. Сниженный мембранный потенциал в диастолический период(50-70мВ)
2. Четвертая фаза не является стабильной и отмечается постепенное снижение мембранного потенциала к пороговому критическому уровню деполяризации и в диастолу постепенно медленно продолжает снижаться достигая критического уровня деполяризации при котором произойдет самовозбуждение П-клеток. В P-клетках происходит усиление проникновения ионов натрия и снижение выхода ионов калия. Повышается проницаемость ионов кальция. Эти сдвиги в ионном составе приводят к тому, что мембранный потенциал в P-клетках снижается до порогового уровня и p-клетка самовозбуждается обеспечивая возникновение потенциала действия. Плохо выражена фаза Плато. Фаза ноль плавно переходи ТВ процесс реполяризации, который восстанавливает диастолический мембранный потенциал, а дальше цикл повторяется вновь и P-клетки переходят в состояние возбуждения. Наибольшой возбудимостью обладают клетки сино-атриального узла. Потенциал в нем особо низок и скорость диастолической деполяризации наиболее высок.. Это будет влиять на частоту возбуждения. P- клетки синусного узла генерируют частоту до 100 ударов в мин. Нервная система(симпатическая система) подавляют действие узла(70 ударов). Симпатическая система может повышать автоматию. Гуморальные факторы- адреналин, норадреналин. Физические факторы – механический фактор – растяжение, стимулируют автоматию, согревание, тоже увеличивает автоматию. Все это применяется в медицине. На этом основано мероприятие прямого и непрямого массажа сердца. Область атриовентрикулярного узла тоже обладает автоматией. Степень автоматии атриовентрикулярного узла выражена значительно меньше и как правило она в 2 раза меньше, чем в синусном узле – 35-40. В проводящей системе желудочков импульсы тоже могут возникать(20-30 в минуту). ПО ходу проводящей системы возникает постипенное снижение уровня автоматии, что получило название градиента автоматии. Синусный узел – центр автоматии первого порядка.
Станеус – ученый. Наложение лигатур на сердце лягушки(3х камерное). У правого предсердия имеется венозныц синус, где лежит аналог синусного узла человека. Станеус накладывал 1ую лигатуру между венозным синусом и предсердием. Когда лигатура затягивалась сердце прекращала свою работу. Вторая лигатура накладывалась Станеусом между предсердиями и желудочком. В этой зоне находится аналог атрии-вентрикулярного узла, но 2ая лигатура имеет задачу не отделения узла, а его механическое возбуждение. Ее накладывают постепенно, возбуждая атриовентрикулярный узел и при этом возникает сокраение сердца. Желудочки получают вновь сокращаться под действием атрии-вентрикулярного узла. С частотой в 2 раза меньше. Если наложить 3ю лигатуру, которая отделяет атривентрикулярный узел возникает остановка сердца. Все это дает нам возможность показать, что синусный узел – водитель ритма, атриовентрикулярный узел обладает меньшей автоматией. В проводящей системе существуе убывающий градиент автоматии.
Физиологические свойства сердечной мышцы.
Возбудимость, проводимость ,сократимость
Под возбудимостью сердечной мышцы понимается ее свойство отвечать на действие раздражителей пороговой или над пороговой силы процессом возбуждения. Возбуждение миокарда можно получить на действие химических, механических, температурных раздражений. Эта способность отвечать на действие разных раздражителей используется при массаже сердца(механическое), введение адреналина, кардиостимуляторы. Особенностью реакции сердца на действие раздражителя, играет то что действует по принципу «Все или ничего». Сердце отвечает максимальным импульсом уже на пороговый раздражитель. Продолжительность сокращения миокарда в желудочкх составляет 0,3с. Это обусловлено длительным потенциалом действия, который тоже длится до 300мс. Возбудимость сердечной мышцы может падать до 0 – абсолютно рефрактерная фаза. Никакие раздражители не могут вызвать повторного возбуждения(0,25-0,27с). Сердечная мышца абсолютно невозбудима. В момент расслабления(диастолы)абсолютная рефрактерная переходит в относительную рефрактерную 0,03-0,05с. В этот момент можно получить повторное раздражение на над пороговые раздражители. Рефрактерный период сердечной мышцы длится и совпадает по времени столько, сколько длится сокращение. Вслед за относительной рефрактерностью имеется небольшой период повышенной возбудимости – возбудимость ставновится выше исходного уровня – супер нормальная возбудимость. В эту фазу сердце особо чувствительно к воздействию других раздражителей(смогут возникать др. раздражители или экстрасистолы- внеочередные систолы). Наличие длительного рефрактерного периода должно оградить сердце от повторных возбуждений. Сердце выполняет насосную функцию. Промежуток между нормальным и внеочередным сокращением укорачивается. Пауза может быть нормальной или удлиненной.Удлиненную паузу называют компенсаторной. Причина экстрасистолов – возникновение других очагов возбуждения – атриовентрикулярный узел, элементы желудочковой части проводящей системы, клетки рабочего миокарда, Это может быть связано с нарушением кровоснабжением, нарушением проведения в сердечной мышцей, но все дополнительные очаги – эктопические очаги возбуждения. В зависимости от локализации – разные экстрасистолы – синусные, предсредные, атриовентрикулярные. Экстрасистолы желудочка сопровождаются удлиненной компенсаторнйо фазой. 3 дополнительное раздражение – причина внеочередного сокращения. Вовремя экстрасистола сердце утрачивает возбудимость. К ним приходит очередной импульс из синусного узла. Пауза нужна для восстановления нормального ритма. Когда в сердце происходит сбой сердце пропускает одно нормальное сокращение и дальше возвращается к нормальному ритму.
Проводимость – способность проводить возбуждение. Скорость проведения возбуждения в разных отделах неодинакова. В миокарде предсердий – 1 м/c и время проведения возбуждения занимает 0,035 с
Скорость проведения возбуждения
Миокард 1 м/c 0,035
A-V узел 0,02 – 0-05 м в с. 0,04 с
Проведение система желудочков – 2-4,2 м в с. 0,32
В сумме от синусного узла до миокарда желудочка – 0,107 с
Миокард желудочка – 0,8-0,9 м в с
Нарушение проведения сердца приводит к развитию блокад – синусной , атривентрикулярной, пучка гиса и его ножек. Синусный узел может выключится.. Включится ли атривентрикулярный узел как водитель ритма? Синусные блокады встречаются редко. Больше в атриовентрикулярных узлах. Удлиение задержки(больше 0,21с) возбуждение доходит до желудочка, хоть и замедленно. Выпадение отдельных возбуждений, которые возникают в синусном узле НАПРИМЕР из 3 доходит только 2 – вторая степень блокады. 3я блокада – предсердия и желудочки работают несогласованно. Блокада ножек и пучка – блокада желудочков. Чаще встречаются блокады ножек пучка Гиса и соответственно желудочек запаздывает за другим.
Сократимость
Кардиомиоциты включают фибриллы, саркомеры. Есть продольные трубочки и Т трубочки наружной мембраны, котоыре входят внутрь на уровне мембраны я. Они широкие. Сократительная фугкция кардиомиоцитов связана с белками миозином и актином. На тонких актиновых белках – система тропонин и тропомиозин. Это не дает головкам миозин сцепляется с головками миозина. Снятие блокировки — ионами кальция. По т трубочкам открываются кальцевые каналы. Повышение кальция в саркоплазме снимает тормозной эффект актина и миозина. Мостики миозина перемещают тонике нити к центру. Миокард подчиняется в сократительной функции 2м законам – все или ничего. Сила сокращения зависит от исходной длины кардиомиоцитов – Франк и Старалинг. Если миоциты предварительно растянуты, то они отвечают большей силой сокращения. Растяжение зависит от наполнения кровью. Чем больше- тем сильней. Этот закон формулируют как – систола есть функция диастолы. Это важный приспособительный механизм. Это синхронизирует работу правого и левого желудочка.
ОСОБЕННОСТИ СВОЙСТВ СЕРДЕЧНОЙ МЫШЦЫ И ЕЕ ЭНЕРГООБЕСПЕЧЕНИЯ
Свойства сердечной мышцы существенно отличаются от свойств скелетной мышцы, что определяется структурными особенностями кардиомиоцитов и их функциональными взаимоотношениями.
А. Возбудимость —
это способность клеток генерировать потенциал действия (ПД).
1. Параметры ПП и ПД.
Потенциал покоя клеток рабочего миокарда формируется в основном градиентом К+, его величина в среднем составляет 85 — 90мВ,
величина ПД 120 мВ. Длительность ПД
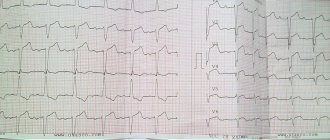
кардиомиоцитов желудочков — 300-400 мс — почти соответствует длительности сокращения мышцы сердца (рис. 8.3). Продолжительность ПД кардиомиоцитов предсердия составляет 100 мс, примерно столько же длится систола предсердий.
2. Ионный механизм возникновения ПД
кардиомиоцитов. Фазы деполяризации и восходящая часть инверсии осуществляются в основном за счет входа №+ в клетку, как у миоцитов скелетной мышцы. В начале нисходящей части фазы инверсии снижение ПД происходит быстро, очевидно, вследствие входа СГ в клетку, согласно концентрационному и электрическому градиентам (отрицательно заряженньшионов хлора, как известно, больше вне клетки, которая во время инверсии внутри заряжена положительно). В кардиомиоцитах обнаружены хлорные потенциалчувствительные каналы. Медленный спад в фазу инверсии (плато) обеспечивается в основном входом Са2+ в клетку. При мембранном потенциале 0 ± 10 мВ, т.е. сразу после фазы инверсии, активация К-каналов быстро нарастает, и К+ начинает лавиной выходить из клетки, что обеспечивает фазу реполяризации кардиомиоцита.
3. Длительность фазы абсолютной рефрактерности кардиомиоцитов в 100-300 раз больше
таковой миоцитов скелетной мышцы. У последней, как известно, пик ПД длится 1-3 мс. Примерно
столько же длится и абсолютная рефрактерная фаза, которая соответствует длительности пика ПД, поскольку во время его пика имеет место инактивация быстрых 1Ма-каналов. Так же и у кардиомиоци-тов — пока длится ПД (около 300 мс), они невозбудимы, что связано в основном с инактивацией быстрых №-каналов: период абсолютной рефрактерности для миокарда желудочков составляет 270 мс; он соответствует фазе инверсии (плато ПД), период относительной рефрактерности — 30 мс — соответствует фазе реполя-ризации (см. рис. 8.3). Длительная рефрактерная фаза предотвращает круговое распространение возбуждения по миокарду, так как время распространения возбуждения меньше длительности рефрактерной фазы.
Б. Проводимость сердечной мышцы принципиально отличается от проводимости скелетной мышцы. В миокарде возбуждение распространяется из любой точки во всех направлениях — диффуз-но (генерализованный характер распространения возбуждения). Это обеспечивается нексусами (тесными контактами) с низким электрическим сопротивлением — они обеспечивают распростра- • нение возбуждения от одного миоцита к другому. Скорость проведения возбуждения по миокарду около 1м/с,что в 3,5 раза меньше, чем у скелетной мышцы.
В. Сократимость сердечной мышцы также существенно отличается от сократимости скелетной мышцы. Во-первых, сердечная мышца, в отличие от скелетной, подчиняется закону «все или ничего»: сердечная мышца либо не отвечает на раздражение, если оно ниже порогового, либо отвечает максимальным сокращением, если раздражитель достигает пороговой или сверхопороговой силы. Увеличение силы раздражения выше пороговой не увеличивает силу сокращения, как при действии на скелетную мышцу. У сердечной мышцы возбуждение, возникнув в одном месте, распространяется диффузно по всем кардиомиоцитам, и все они вовлекаются в сокращение. Во-вторых, у сердечной мышцы более длительный период одиночного сокращения —
он примерно соответствует длительности ПД.
В-третьих, сердечная мышца не может сокращаться тетанически. Это объясняется тем, что у нее длительный рефрактерный период — он равен продолжительности ПД и одиночного сокращения мышцы. Это означает, что весь период сокращения мышца сердца невозбудима. Возникновение тетанического сокращения могло бы нарушить нагнетательную функцию сердца.
Г. Растяжимость и эластичность сердечной мышцы также играют важную роль в выполнении нагнетательной функции сердца. Во-первых, оба этих свойства смягчают гидравлический удар, возникающий в результате несжимаемости жидкости и быстро сокращающихся стенок сердца. Во-вторых, эластические силы, возникающие вследствие растяжения стенок сердца при его наполнении кровью, в том числе и при сокращении предсердий, увеличивают силу сокращений миокарда в начале систолы. В-третьих, эластичность структурных элементов сердца обеспечивает возникновение в конце систолы упругих сил, способствующих расслаблению сократившегося (сжавшегося) миокарда после прекращения его возбуждения. Стенки (особенно желудочков) расслабляются — расправляются, как предварительно сдавленная резиновая груша, так как различное направление отдельных мышечных слоев друг относительно друга увеличивает упругость стенок сердца, которая превышает упругость скелетной мышцы.
Д. Главным источником энергии для сердца является процесс аэробного окисления. Анаэробное окисление
(анаэробный гликолиз) для сердца, в отличие от скелетной мышцы, играет незначительную роль. Источником энергии в сердце, благодаря аэробному окислению, являются главным образом неуглеводные субстраты. Это свободные жирные кислоты и молочная кислота (около 60%), пировиноградная кислота, кетоновые тела и аминокислоты (менее 10%). При интенсивной мышечной работе в крови накапливается молочная кислота в результате анаэробного гликолиза в мышцах. Лактат является дополнительным источником энергии для сердца, причем, расщепляя молочную кислоту, сердце способствует поддержанию постоянства рН. Только около 30% расходуемой сердцем энергии покрывается за счет глюкозы; при физической нагрузке увеличивается энергетическая доля жирных и молочной кислот при одновременном снижении энергетической доли глюкозы. Таким образом, сердце утилизирует недоокисленные продукты, накапливающиеся в результате интенсивной мышечной работы, и тем самым препятствует закислению внутренней среды организма.
Большая зависимость деятельности сердечной мышцы от аэробного окисления делает сердце весьма зависимым от по- . ступления кислорода
к кардиомиоцитам. Сердце на 1 кг массы потребляет 02 в 25 раз больше. Скелетная мышца может некоторое время работать вообще без кислорода (в долг) за счет гликолиза. Коэффициент полезного действия сердца в среднем составляет около 30%, то есть несколько больше, нежели скелетной мышцы -ее КПД 20-25%.
При ухудшении коронарного
кровотока и недостаточном поступлении кислорода к сердечной мышце в ней могут развиваться патологические процессы, вплоть до инфаркта. Однако это случается относительно редко благодаря
миоглобину, имеющемуся в сердечной мышце в количестве около 4 мг/г ткани. Он обладает большим сродством к 02, запасает его во время диастолы и покоя сердца и отдает во время систолы, когда кровоток в коронарных артериях желудочков прекращается.
Свойства сердечной мышцы существенно отличаются от свойств скелетной мышцы, что определяется структурными особенностями кардиомиоцитов и их функциональными взаимоотношениями.
А. Возбудимость —
это способность клеток генерировать потенциал действия (ПД).
1. Параметры ПП и ПД.
Потенциал покоя клеток рабочего миокарда формируется в основном градиентом К+, его величина в среднем составляет 85 — 90мВ,
величина ПД 120 мВ. Длительность ПД
кардиомиоцитов желудочков — 300-400 мс — почти соответствует длительности сокращения мышцы сердца (рис. 8.3). Продолжительность ПД кардиомиоцитов предсердия составляет 100 мс, примерно столько же длится систола предсердий.
2. Ионный механизм возникновения ПД
кардиомиоцитов. Фазы деполяризации и восходящая часть инверсии осуществляются в основном за счет входа №+ в клетку, как у миоцитов скелетной мышцы. В начале нисходящей части фазы инверсии снижение ПД происходит быстро, очевидно, вследствие входа СГ в клетку, согласно концентрационному и электрическому градиентам (отрицательно заряженньшионов хлора, как известно, больше вне клетки, которая во время инверсии внутри заряжена положительно). В кардиомиоцитах обнаружены хлорные потенциалчувствительные каналы. Медленный спад в фазу инверсии (плато) обеспечивается в основном входом Са2+ в клетку. При мембранном потенциале 0 ± 10 мВ, т.е. сразу после фазы инверсии, активация К-каналов быстро нарастает, и К+ начинает лавиной выходить из клетки, что обеспечивает фазу реполяризации кардиомиоцита.
3. Длительность фазы абсолютной рефрактерности кардиомиоцитов в 100-300 раз больше
таковой миоцитов скелетной мышцы. У последней, как известно, пик ПД длится 1-3 мс. Примерно
столько же длится и абсолютная рефрактерная фаза, которая соответствует длительности пика ПД, поскольку во время его пика имеет место инактивация быстрых 1Ма-каналов. Так же и у кардиомиоци-тов — пока длится ПД (около 300 мс), они невозбудимы, что связано в основном с инактивацией быстрых №-каналов: период абсолютной рефрактерности для миокарда желудочков составляет 270 мс; он соответствует фазе инверсии (плато ПД), период относительной рефрактерности — 30 мс — соответствует фазе реполя-ризации (см. рис. 8.3). Длительная рефрактерная фаза предотвращает круговое распространение возбуждения по миокарду, так как время распространения возбуждения меньше длительности рефрактерной фазы.
Б. Проводимость сердечной мышцы принципиально отличается от проводимости скелетной мышцы. В миокарде возбуждение распространяется из любой точки во всех направлениях — диффуз-но (генерализованный характер распространения возбуждения). Это обеспечивается нексусами (тесными контактами) с низким электрическим сопротивлением — они обеспечивают распростра- • нение возбуждения от одного миоцита к другому. Скорость проведения возбуждения по миокарду около 1м/с,что в 3,5 раза меньше, чем у скелетной мышцы.
В. Сократимость сердечной мышцы также существенно отличается от сократимости скелетной мышцы. Во-первых, сердечная мышца, в отличие от скелетной, подчиняется закону «все или ничего»: сердечная мышца либо не отвечает на раздражение, если оно ниже порогового, либо отвечает максимальным сокращением, если раздражитель достигает пороговой или сверхопороговой силы. Увеличение силы раздражения выше пороговой не увеличивает силу сокращения, как при действии на скелетную мышцу. У сердечной мышцы возбуждение, возникнув в одном месте, распространяется диффузно по всем кардиомиоцитам, и все они вовлекаются в сокращение. Во-вторых, у сердечной мышцы более длительный период одиночного сокращения —
он примерно соответствует длительности ПД.
В-третьих, сердечная мышца не может сокращаться тетанически. Это объясняется тем, что у нее длительный рефрактерный период — он равен продолжительности ПД и одиночного сокращения мышцы. Это означает, что весь период сокращения мышца сердца невозбудима. Возникновение тетанического сокращения могло бы нарушить нагнетательную функцию сердца.
Г. Растяжимость и эластичность сердечной мышцы также играют важную роль в выполнении нагнетательной функции сердца. Во-первых, оба этих свойства смягчают гидравлический удар, возникающий в результате несжимаемости жидкости и быстро сокращающихся стенок сердца. Во-вторых, эластические силы, возникающие вследствие растяжения стенок сердца при его наполнении кровью, в том числе и при сокращении предсердий, увеличивают силу сокращений миокарда в начале систолы. В-третьих, эластичность структурных элементов сердца обеспечивает возникновение в конце систолы упругих сил, способствующих расслаблению сократившегося (сжавшегося) миокарда после прекращения его возбуждения. Стенки (особенно желудочков) расслабляются — расправляются, как предварительно сдавленная резиновая груша, так как различное направление отдельных мышечных слоев друг относительно друга увеличивает упругость стенок сердца, которая превышает упругость скелетной мышцы.
Д. Главным источником энергии для сердца является процесс аэробного окисления. Анаэробное окисление
(анаэробный гликолиз) для сердца, в отличие от скелетной мышцы, играет незначительную роль. Источником энергии в сердце, благодаря аэробному окислению, являются главным образом неуглеводные субстраты. Это свободные жирные кислоты и молочная кислота (около 60%), пировиноградная кислота, кетоновые тела и аминокислоты (менее 10%). При интенсивной мышечной работе в крови накапливается молочная кислота в результате анаэробного гликолиза в мышцах. Лактат является дополнительным источником энергии для сердца, причем, расщепляя молочную кислоту, сердце способствует поддержанию постоянства рН. Только около 30% расходуемой сердцем энергии покрывается за счет глюкозы; при физической нагрузке увеличивается энергетическая доля жирных и молочной кислот при одновременном снижении энергетической доли глюкозы. Таким образом, сердце утилизирует недоокисленные продукты, накапливающиеся в результате интенсивной мышечной работы, и тем самым препятствует закислению внутренней среды организма.
Большая зависимость деятельности сердечной мышцы от аэробного окисления делает сердце весьма зависимым от по- . ступления кислорода
к кардиомиоцитам. Сердце на 1 кг массы потребляет 02 в 25 раз больше. Скелетная мышца может некоторое время работать вообще без кислорода (в долг) за счет гликолиза. Коэффициент полезного действия сердца в среднем составляет около 30%, то есть несколько больше, нежели скелетной мышцы -ее КПД 20-25%.
При ухудшении коронарного
кровотока и недостаточном поступлении кислорода к сердечной мышце в ней могут развиваться патологические процессы, вплоть до инфаркта. Однако это случается относительно редко благодаря
миоглобину, имеющемуся в сердечной мышце в количестве около 4 мг/г ткани. Он обладает большим сродством к 02, запасает его во время диастолы и покоя сердца и отдает во время систолы, когда кровоток в коронарных артериях желудочков прекращается.